ЭКСПРЕСС-ОЦЕНКА ОБРАЗЦОВ РИСА НА ПРИСУТСТВИЕ ГЕНА УСТОЙЧИВОСТИ К ЗАТОПЛЕНИЮ SUB1A
Научная статья
Костылев П.И.1, *, Галаян А.Г.2, Казакова А.С.3, Костылева Л.М.4
1 ORCID: 000-0002-4371-6848,
1 ФГБНУ «АНЦ «Донской»;
2 ORCID: 0000-0003-4174-9731;
3 ORCID: 0000-0002-0957-3994;
4 ORCID: 0000-0003-3078-2296;
2, 3, 4 Азово-Черноморский инженерный институт ФГБОУ ВО Донской ГАУ, Зерноград, Россия
* Корреспондирующий автор (p-kostylev[at]mail.ru)
Аннотация
В статье представлены результаты исследований методических аспектов экспресс-оценки линий риса по их реакции на условия аэробного и анаэробного прорастания в лабораторных условиях. Изучены линии шестого поколения гибридов от скрещивания устойчивых к длительному затоплению сортов с геном Sub1A: BR-11, CR-1009, Inbara 3, TDK-1 с отечественным сортом Новатор, полученные в АНЦ «Донской». Семена проращивали в пробирках, залитых водой и чашках Петри. Установлено, что образцы, не растущие под водой, несли ген Sub1A в гомо- и гетерозиготном состоянии, а у растущих проростков он отсутствовал. Следовательно, данную методику можно использовать при быстрой и экономически выгодной оценке гибридного материала.
Ключевые слова: рис, линия, устойчивость к затоплению, экспресс-оценка, ген Sub1A.
EXPRESS EVALUATION OF RICE SAMPLES FOR IDENTIFICATION OF THE GENE RESISTANT TO FLOOD SUB1A
Research article
Kostylev P.I.1, *, Galayan A.G.2, Kazakova A.S.3, Kostyleva L.M.4
1 ORCID: 0000-0002-4371-6848,
1 FSBSI “”ANC” Donskoy”;
2 ORCID: 0000-0003-4174-9731;
3 ORCID: 0000-0002-0957-3994;
4 ORCID: 0000-0003-3078-2296;
2, 3, 4 Azov-Black Sea Engineering Institute of FSBEI HE Donskoy SAU, Zernograd, Russia
* Corresponding author (p-kostylev[at]mail.ru)
Abstract
The article presents the results of research on the methodological aspects of rapid assessment of rice lines by their reaction to the conditions of the laboratory aerobic and anaerobic germination. The lines of the sixth generation of hybrids from crossing resistant to long-term flooding of varieties with the Sub1A gene: BR-11, CR-1009, Inbara 3, TDK-1 with the domestic variety Novator obtained in the ANC “Donskoy” were studied. Seeds were germinated in test tubes filled with water and Petri dishes. It was established that the samples that did not grow under water carried the Sub1A gene in the homo- and heterozygous state, while it was absent in growing seedlings. This technique can be subsequently used in a quick and cost-effective assessment of the hybrid material.
Keywords: rice, line, resistance to flooding, rapid assessment, Sub1A gene.
Во всей Юго-Восточной Азии ежегодные внезапные наводнения и тайфуны приводят к потерям урожая риса. Только в Бангладеш и Индии они составляют до 4 миллионов тонн риса в год – этого достаточно, чтобы прокормить 30 миллионов человек. Длительное погружение в воду является третьим по значимости абиотическим стрессом, сдерживающим продуктивность риса. Хотя рис является культурой, которая требует затопленных и орошаемых условий для выращивания, большинство сортов риса подвержены наводнениям, и если вода застаивается, удерживая растения погруженными в воду более, чем на семь дней, это вызывает удлинение листьев и стеблей, гниение листьев, потерю сухой массы, а также полегание после сброса воды [1. С. 899].
При затоплении длительностью в семь дней погибает большинство растений риса, но некоторые растения риса могут находиться в состоянии покоя под водой в течение двух недель.
Толерантность к длительному затоплению была открыта у индийского сорта FR13A, именно это привело к успешной селекции устойчивых сортов. При длительном глубоком затоплении выживают генотипы, сочетающие экономию энергии и углеводов, контролируемую геном Sub 1. Этот механизм выражается в состоянии покоя и отсутствии удлинения проростков под водой [2, С.1107].
Идентификация на 9-й хромосоме гена Sub1, контролирующего толерантность к погружению, позволила создать в Международном институте риса (IRRI) с помощью беккроссов новые высокоурожайные коммерческие «мегаcорта» с такими важными характеристиками, как высокая урожайность, хорошее качество зерна и устойчивость к местным вредителям и болезням [3, С. 227].
Толерантность к затоплению или анаэробная толерантность характеризуется способностью растений восстановиться после затопления на разных этапах развития. Восстановление включает в себя регенерацию растений, поврежденных внезапным затоплением [4, С. 170]. Основания для толерантности к затоплению связанны с высоким начальным содержанием углеводов, высоким уровнем хлорофилла и минимальным ростом в условиях погружения [5, С. 77], [6, С. 2030]. Толерантность к затоплению генетическое свойство, которое проявляется в условиях стресса из-за участия многих неконтролируемых факторов окружающей среды. Xu и Mackill [7, С. 222] разработали метод регенерации проростков для оценки толерантности при затоплении риса, а Yamauchi и Winn [8, С. 683] разработали метод появления всходов для экспресс-оценки всходов при затоплении. При наличии большого количества генетических ресурсов риса из них можно выбрать перспективные генотипы с желательными признаками.
Цель исследований – разработка простого, быстрого и надежного способа экспресс-оценки генотипов риса с высокой устойчивостью проростков к условиям затопления.
Материал и методика
В качестве селекционного материала использовали 64 линии шестого поколения сорта из гибридных комбинаций: BR-11 x Новатор, CR-1009 x Новатор, Inbara 3 x Новатор, TDK-1 x Новатор, полученные в АНЦ «Донской». Материнские позднеспелые родительские формы несли ген Sub1A, отцовский краснодарский раннеспелый сорт Новатор – нет. Ген устойчивости к длительному затоплению Sub1A выявляли с помощью ПЦР-анализа растений по маркеру доминантного аллеля RM7481.
Опытным вариантом оценки устойчивости риса к длительному затоплению было анаэробное прорастание. Семена (3 шт.) помещали в стеклянную пробирку (2,5 см в диаметре и 15 см в высоту), заливали дистиллированной водой глубиной 10 см и инкубировали при температуре 28°С в темноте, не меняя воду. На 15-й день измеряли длину проростка (ростка и корешка).
Контрольным вариантом было аэробное прорастание на влажной фильтровальной бумаге в чашках Петри при температуре 28°С в темноте. Через 7 суток проростки измеряли и помещали в пробирки под слой воды 10 см при температуре 28°С. На 7-й день инкубации измеряли длину проростка и сравнивали с начальной величиной [9, С. 268]. Статистическую обработку полученных данных проводили с помощью программы Excel и Statistica 6.
Результаты исследований
В результате исследований было установлено, что при анаэробном прорастании через 15 суток большинство изучаемых сортов и линий (63 %) имели среднюю длину ростка от 0,5 до 2,5 см, только 11 % обладали максимальной средней длиной проростков от 3,0 до 4,5 см (рис. 1).
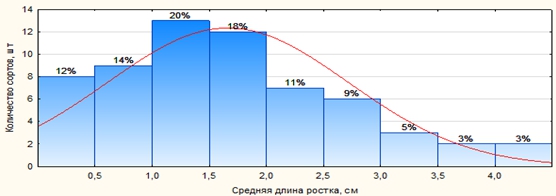
Рис. 1 – Распределение сортов и линий риса по средней длине проростка при анаэробном прорастании
Максимальное значение средней длины ростка в интервале от 3,10 до 4,50 см было у линий 6469 (4,49 см), 6502 (4,28 см), 6521 (3,98 см), 6465 (3,64 см), 6552 (3,42 см), 6446 (3,20 см), 6528 (3,10 см).
Большинство линий не имели корней. Только у трех линий и одного сорта был обнаружен проросший корешок, его длина составила у линии 6592 – 0,7 см, 6506 – 0,6 см, 6474 – 0,5 см, у сорта Южанин – 0,45 см.
R. Lasanthi-Kudahettige иL. Magneschi (2007) утверждают, что при аноксии проросток риса удлиняет междоузлие и формирует в первую очередь росток, более длинный, чем при аэробном прорастании, чем и обуславливается незначительное развитие или отсутствие корешка у большинства растений риса [10]. В процессе исследования три линии риса (6505, 6484 и 6529) не проросли в пробирках, что первоначально не дало достоверной информации о наличии гена Sub 1A, так как не исключается нежизнеспособность семенного материала при его проращивании. Однако при проращивании в чашках Петри все образцы проросли, следовательно, отсутствие прорастания в пробирках обусловлено присутствием в их генотипах гена Sub 1A..
Анализ аэробного прорастания в чашках Петри показал, что на 3 день закладки опыта, максимальные значения средней длины ростка в интервале от 0,14 до 0,98 см были у линий 6461 (0,98 см), 6517 (0,77 см), величина средней длины корешка варьировала в интервале от 0,44 до 1,98 см, максимум – у линий 6461 (1,98 см), 6543 (1,62 см). На 8 день закладки опыта: средняя длина ростка колебалась от 0,26 до 1,03 см (максимум у линии 6469 – 1,03 см), средняя длина корешка варьировала от 0,27 до 1,98 см (у линии 6461 – 1,98 см.
По данным изменения величины прироста образцов можно судить о наличии или отсутствии гена Sub 1A. Через неделю после переноса проростков из чашек Петри в пробирки у 24 образцов и сортов увеличение длины проростка варьировало от 0,1 до 0,4 см (максимум у линии 6549). У остальных 41 образцов прироста не наблюдалось. Минимальный прирост, или состояние покоя у изучаемых образцов свидетельствует о возможности присутствия в них гена Sub1A.
Поэтому был проведен анализ ДНК-маркеров этого гена. По данным ПЦР-анализа, изученные линии распределились на 3 группы по наличию гена Sub 1А: гомозиготы по рецессивному аллелю (нижний бэнд), гомозиготы по доминантному аллелю (верхний бэнд) и гетерозиготы (оба бэнда) (рис. 2).
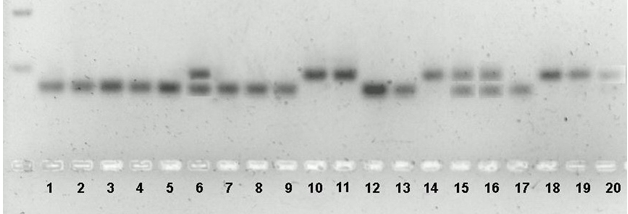
Рис. 2 – Электрофореграмма анализа наличия гена Sub1A
Примечание: Рецессивные гомозиготы: 1 – 6417, 2 – 6436, 3 – 6437, 4 – 6469, 5 – 6504, 7 – 6513, 8 – 6530, 9 – 6545, 12 – Новатор (реципиент), 13 – 6549, 17 – 6466, доминантные гомозиготы Sub1A: 10 – 6429, 11 – BR-11 (донор), 14 – 6461, 18 – 6484, 19 – 6505, 20 – 6529, гетерозиготы: 6 – 6477, 15 – 6546, 16 – 6592
Оказалось, что образцы без прироста под водой несли ген Sub1A в гомо- и гетерозиготном состоянии, а у растущих далее он отсутствовал. Следовательно, данную методику можно использовать при быстрой оценке гибридного материала. Затраты на 1 образец при проведении анализа наличия гена Sub 1А в полевых условиях составляют 300 рублей, с помощью ПЦР-анализа – 200, а при экспресс-оценке – 80.
Таким образом, представленный в данной статье метод является не только простым, быстрым, но и экономически выгодным в использовании по сравнению с другими методами.
| Конфликт интересов
Не указан. |
Conflict of Interest
None declared. |
Список литературы / References
- Scuba rice: breeding flood-tolerance into Asia’s local rice varieties, 2010 // https://assets.publishing.service.gov.uk/media/57a08b0d40f0b652dd000a80/DFID_impact_case_study_SUB1rice_FINAL_1_.pdf site visited on ../0../2019
- Kende H. Rice: a model plant to study stem elongation / H. Kende //Plant Physiol. – 1998. – 118 – P. 1105–1110.
- Submergence tolerance in rainfed lowland rice: Physiological basis and prospects for cultivar improvement through marker–aided breeding // https://www.scilit.net/article/328b8a343d143d4326d381c5f0ea7636 site visited on 04/05/2019
- Kamolsukyunyong W. Mapping of quantitative trait locus related to submergence tolerance in rice with aid of chromosome walking / W. Kamolsukyunyong, V. Ruanjaichon // DNA Res. – 2001. – 8. – P.163–171.
- Ito O. Physiological basis of submergence tolerance in rainfed lowland rice ecosystem / O. Ito, E. Ella, N. Kawano // Field Crops Res. –1999. – 64 – P.75–90.
- Takeshi F. A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice. / F. Takeshi, K. Xu, P. C. Ronald, J. Bailey-Serres // Plant Cell. – 2006. – 18. – P. 2021–2034.
- Xu K. A major locus for submergence tolerance mapped on rice chromosome 9 / K. Xu, D. Mackill // Mol. Breed. – 1996. – 2. – P.219–224.
- Yamauchi M. Rice seed vigor and seedling establishment in anaerobic soil / M. Yamauchi, T. Winn // Crop Sci. – 1996. – 36. – P. 680–686.
- Manangkil, O.E. A simple, rapid and reliable bioassay for evaluating seedling vigor under submergence in indica and japonica rice (Oryza sativa) / O.E. Manangkil, H.T.T. Vu, S. Yoshida, N. Mori, C. Nakamura // Euphytica. – 2008. – V.163. – I.2. – Р. 267-274.
- Transcript Profiling of the Anoxic Rice Coleoptile // http://www. plantphysiol.org /content/144/1/218 site visited on 3/05/2019