СТРЕССОУСТОЙЧИВОСТЬ ИНБРЕДНЫХ ЛИНИЙ DROSOPHILA MELANOGASTER И DROSOPHILA FUNEBRIS С ПОВЫШЕННОЙ ПРОДОЛЖИТЕЛЬНОСТЬЮ ЖИЗНИ
Научная статья
Булыгин А.А.1, *, Пастухова Е.И.2
1 ORCID: 0000-0002-6689-4647;
2 ORCID: 0000-0003-4107-5669;
1, 2 ФГБОУ ВО Челябинский государственный университет, Челябинск, Россия
* Корреспондирующий автор (skytolya[at]ya.ru)
Аннотация
В данной работе были изучены 14 инбредных линий D. melanogaster и D. funebris, выведенных из природных популяций и доведённых до 12-го поколения. Среди инбредных обнаружилось несколько линий с повышенной продолжительностью жизни. Эти линии затем были дополнительно протестированы на стрессоустойчивость тепловым (39⁰C) и холодовым (-1⁰C) шоком и на изменение массы тела относительно мух диких популяций. В итоге выяснилось, что две линии обладают свойствами, способствующими повышенной продолжительности жизни.
Ключевые слова: инбридинг, инбредная депрессия, долгожительство, стрессоустойчивость, плодовая муха.
STRESS RESISTANCE OF INBRED LINES OF DROSOPHILA MELANOGASTER AND DROSOPHILA FUNEBRIS WITH INCREASED LIFE EXPECTANCY
Research article
Bulygin A.A.1, *, Pastukhova E.I.2
1 ORCID: 0000-0002-6689-4647;
2 ORCID: 0000-0003-4107-5669;
1, 2 FSBEI HE Chelyabinsk State University, Chelyabinsk, Russia
* Corresponding author (skytolya[at]ya.ru)
Abstract
In this study, 14 inbred lines of D. melanogaster and D. funebris, derived from natural populations and brought to the 12th generation, were studied. Several lines with increased life expectancy were found among the inbred. These lines were further tested for stress resistance with thermal (39⁰C) and cold (-1⁰C) shock and for changes in body weight relative to flies of wild populations. As a result, it turned out that the two lines have properties that contribute to increased life expectancy.
Keywords: inbreeding, inbreeding depression, longevity, resistance to stress, fruit fly.
Введение
В настоящее время актуальным вопросом науки является изучение биологических механизмов и причин старения организмов. Одним из важнейших объектов биологии и генетики является Drosophila melanogaster. Выявлены различные гены, способные при мутировании существенно изменять продолжительность жизни этого вида [1]. Актуальным вопросом является обнаружение новых генов и сочетаний аллелей, оказывающих влияние на жизнеспособность и продолжительность жизни.
При проведении инбридинга у аутбредных особей могут наблюдаться противоположно направленные тенденции касательно их жизнеспособности и продолжительности жизни: 1) с одной стороны, частым последствием инбридинга является инбредная депрессия, происходящая из-за выхода в гомозиготу рецессивных мутаций или из-за снижения общей гетерозиготности генотипа; 2) с другой стороны в ряде экспериментов обнаружена бо́льшая жизнеспособность потомков нескольких поколений инбридинга [2]. В нашей лаборатории была однажды получена инбредная линия D. funebris с повышенной продолжительностью жизни. Мы решили провести более масштабное исследование по этому поводу. Описанные в этой статье опыты являются продолжением опытов по исследованию инбредных линий из нашей предыдущей статьи [3].
Методы и принципы исследования
Инбредные линии были созданы на основе мух диких популяций, содержащихся в лаборатории кафедры радиационной биологии ЧелГУ на протяжении трёх лет. Для получения нового инбредного поколения самца и самку, отсаживали в пробирку объёмом 15 мл с 5 мл питательной среды стандартного состава [4], смазанной дрожжами. Наркотизация мух проводилась с помощью диэтилового эфира. Инбредные линии удалось вывести с коэффициентом инбридинга от 0,8 до 0,9.
Для опытов на продолжительность жизни девственных мух каждого пола сажали по 5 штук в пробирки того же объёма. Пересадка на новую среду происходила два раза в неделю. D. melanogaster содержались в термостате при постоянном свете и температуре 21⁰C. D. funebris содержались в комнате при температуре 24-25⁰C. Во всех остальных опытах температура находилась в промежутке 24-25⁰C.
В опыте по измерению массы 3-дневные самцы и 5-дневные самки кучками по 8 штук взвешивались на весах с точностью в 0,0001 г.
Для опыта на стрессоустойчивость использовались методы теплового (39⁰C) и холодового (-1⁰C) шока. Во время опыта при 39⁰C записывались времена нокдауна. После термостата или холодильника мух пересыпали в новые пробирки и наблюдали их восстановление. Через сутки считали долю выживших.
Статистическая обработка данных была проведена в программе Past 3.16. Для сравнения кривых выживаемости Каплана-Мейера использовался лог-ранговый тест. Для попарных сравнений масс кучек мух, а также попарных сравнений кривых нокдауна и восстановления использовался U-критерий с поправкой Бонферрони. Различия считались статистически значимыми при p-значении меньшем 0,01.
Основные результаты
По результатам лог-рангового теста кривых выживаемости Каплана-Мейера (табл. 1) для последующих опытов в качестве долгожителей были выбраны линии 3 и 7 у D. melanogaster и 3, 5, 7, 8 у D. funebris.
Таблица 1 – Результаты опыта на продолжительность жизни
| D. melanogaster | ||||||||
| Линия | контроль | 1 | 2 | 3 | 6 | 7 | 8 | 9 |
| Самцы | ||||||||
| Средняя ПЖ, дни | 58 | 50 | 41 | 74 | 59 | 73 | 39 | 64 |
| р | – | < 10-8 | < 10-10 | < 0,05 | > 0,05 | < 0,01 | < 10-10 | > 0,05 |
| Самки | ||||||||
| Средняя ПЖ, дни | 51 | 43 | 44 | 59 | 46 | 57 | 51 | 57 |
| р | – | < 10-3 | < 0,05 | > 0,05 | > 0,05 | > 0,05 | > 0,05 | > 0,05 |
| D. funebris | ||||||||
| Линия | контроль | 1 | 3 | 5 | 6 | 7 | 8 | 9 |
| Самцы | ||||||||
| Средняя ПЖ, дни | 37 | 23 | 64 | 47 | 27 | 39 | 46 | 31 |
| р | – | < 10-14 | < 10-15 | < 10-6 | < 10-8 | > 0,05 | < 10-3 | < 0,05 |
| Самки | ||||||||
| Средняя ПЖ, дни | 39 | 30 | 60 | 54 | 27 | 53 | 57 | 39 |
| р | – | < 10-7 | < 10-9 | < 10-9 | < 10-10 | < 10-6 | < 10-9 | > 0,05 |
Результаты взвешивания мух долгоживущих линий приведены на рисунке 1 (самцы отмечены голубым цветом, самки – розовым, звёздочками отмечены статистически значимые отличия). Единственной линией, масса мух которой значительно ниже контроля, оказалась №3 D. funebris. Линии D. melanogaster не отличились одновременным снижением массы самцов и самок.
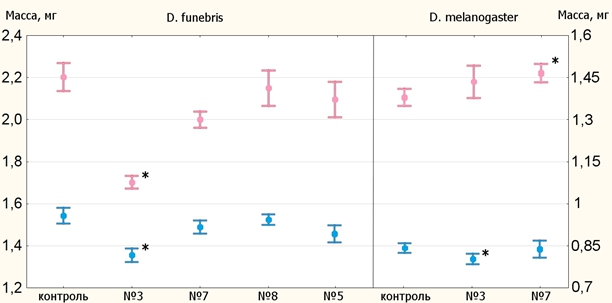
Рис. 1 – Средняя масса мух долгоживущих линий и 95%-ые доверительные интервалы
По результатам опыта на устойчивость к тепловому шоку можно выделить линию №5 D. funebris (рис. 2, 3): самцы этой линии оказались не хуже контроля во времени нокдауна, а самки – лучше контроля как по времени нокдауна, так и по времени восстановления. Опыт по определению времени восстановления самцов D. funebris не удался.
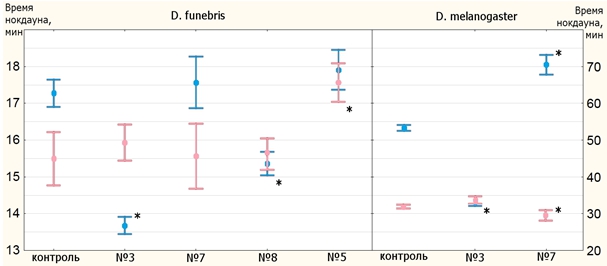
Рис. 2 – Средние времена нокдауна (39⁰C)
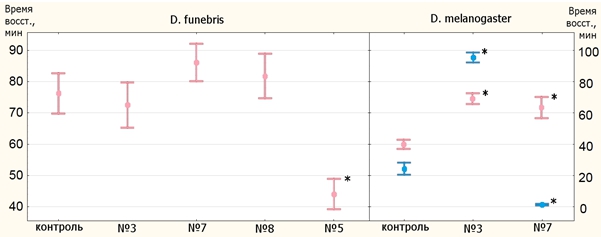
Рис. 3 – Средние времена восстановления (39⁰C)
В опыте на устойчивость к холодовому шоку были проверены только линии D. funebris (рис. 4, табл. 2). Мухи находились при температуре -1⁰C 4 часа. В силу слишком малого времени нокдауна различия между линиями могли быть определены лишь по времени восстановления. Ни одна линия не показали результаты лучше, чем у контроля. Пятая линия оказалась на уровне контроля. Диаграмма для самок выглядит очень похоже на диаграмму для самцов, поэтому на рисунке не изображена.
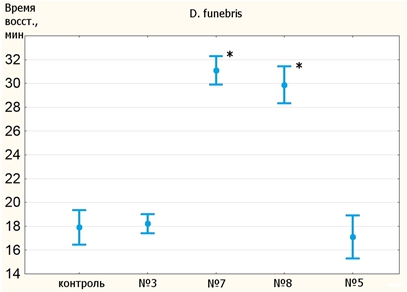
Рис. 4 – Средние времена восстановления самцов линий D. funebris (-1⁰C)
Таблица 2 – Показатели восстановления D. funebris после холодового шока
| Самцы | |||||
| Линия | контроль | 3 | 7 | 8 | 5 |
| Среднее время восст., мин | 17,9 | 18,2 | 31,1 | 30 | 17,1 |
| p | – | 1 | <10-17 | <10-11 | 1 |
| Выживаемость | 0,72 | 0,74 | 0,06 | 0,79 | 0,98 |
| Самки | |||||
| Линия | контроль | 3 | 7 | 8 | 5 |
| Среднее время восст., мин | 17,8 | 18,3 | 29,9 | 29 | 17,6 |
| p | – | 1 | <10-17 | <10-10 | 1 |
| Выживаемость | 0,68 | 0,64 | 0,09 | 0,71 | 0,98 |
Обсуждение
Продолжительность жизни инбредных мух была исследована в очень большом количестве работ. Лучшими, кого удавалось получить в них, были инбредные линии, продолжительность жизни которых была на уровне аутбредных. Такие результаты были получены, например, в работе [5]. В той же работе было показано, что чем ниже температура содержания мух в опыте на продолжительность жизни, тем больше различия между кривыми выживаемости. При переходе от 25⁰C к 21⁰C разница между медианами ПЖ инбредных и аутбредных линий увеличилась в среднем в 1,5 раза. Можно предположить, что в нашем случае выбор температуры 25⁰C вместо 21⁰C для D. melanogaster привёл бы к отсутствию статистически значимых отличий для линий 3 и 7. Но, с другой стороны, возможно, именно пониженная температура позволила их заметить.
Неожиданным результатом является статистически значимое (р<10-4) увеличение ПЖ самцов линий 3 и 7 D. melanogaster относительно самок, при том, что мухи контрольной линии живут примерно одинаковое время. Это не объясняется стандартными гипотезами о влиянии инбридинга на продолжительность жизни. Во всех изученных нами работах ПЖ самцов инбредных линий не была выше ПЖ самок (например, [6]). Для D. funebris существенных противоречий не наблюдается: контрольные самки и самцы живут примерно одинаковое время, а самки долгоживущих линий живут дольше самцов (p<0,01).
Опубликованных работ по исследованию массы инбредных мух нами найдено не было. В работах по отбору мух по массе тела корреляций с продолжительностью жизни не обнаруживали [7]. Корреляции были обнаружены в опытах по отбору в условиях перенаселения [4]. Но авторы данной работы показывают, что снижение массы и увеличение ПЖ оба являются следствием отбора и не являются причиной одно другого.
Снижение массы тела, а точнее снижение количества клеток в организме, является косвенной причиной увеличения продолжительности жизни при нарушениях инсулинового сигналинга. Речь идёт о пути dInR/dPI3K/dAKT, из-за подавления некоторых генов которого клетки начинают реже делиться. Значительное увеличение ПЖ вызывает, например, подавление гена 14-3-3ε, отрицательного регулятора dFoxO, который сам ингибирует клеточное деление при наличии инсулина [8]. В нашей работе не было никакого отбора по названным выше критериям, поэтому можно предположить, что увеличение ПЖ линии №3 D. funebris сверх нормального уровня произошло из-за избегания инбредной депрессии и дополнительно появления и/или закрепления какой-либо мутации, нарушающей инсулиновый сигналинг.
В исследованиях струссоустойчивости инбредных мух обнаруживали только отрицательное действие инбридинга. Но в работе [9] было получено, что при инбредной депрессии значительное изменение экспрессии в ответ на стресс происходит не более, чем у 200 генов из почти 10 тысяч исследованных. Авторы пришли к выводу, что это подтверждает гипотезу о первоочерёдной роли в инбредной депрессии определённых рецессивных аллелей, а не повышенной гомозиготности генома в целом.
Низкую продолжительность жизни инбредных линий можно восстановить до нормального уровня, изменив экспрессию генов, связанных с ответом на стресс. Такой результат был получен, например, для гена Indy [10]. Для этого же гена показано и увеличение ПЖ сверх нормального уровня. Сверхэкспрессия супероксиддисмутазы приводит лишь к восстановлению ПЖ до нормального уровня у инбредных линий, но не превышению его [11].
Подобные результаты получали и в работах по отбору мух по ПЖ. В работе [12] обнаружили улучшение экспрессии некоторых генов, связанных с ответом на стресс (mthl, Or83b, Gr64a и др). В работе по отбору мух по стрессоустойчивости [13] авторы получили долгоживущих мух 37-го поколения отбора.
Соотнося результаты описанных исследований и наши, мы предполагаем, что линия №5 D. funebris смогла избежать инбредной депрессии и превысила нормальную ПЖ за счёт лучшей стрессоустойчивости. Относительно линии №7 D. melanogaster, самцы которой показали повышенную стрессоустойчивость, а самки – пониженную [3], мы не можем ничего сказать однозначно. Удивительным результатом является и линия №8 D. funebris, показавшая одну из лучших ПЖ, и имеющая одну из худших стрессоустойчивость при нормальной массе. Линии №3 D. melanogaster и №7 D. funebris, имеющие ПЖ, немногим выше контроля, плохие показатели стрессоустойчивости и нормальную массу, скорее всего имеют инбредную депрессию.
Заключение
В данной работе мы подвели итоги нашего исследования инбредных линий D. melanogaster и D. funebris. Из шести инбредных линий с повышенной продолжительностью жизни у двух наблюдалось улучшение показателей, способствующих этому повышению: линия 3 D. funebris отличилась сниженной массой тела, линия 5 D. funebris – повышенной устойчивостью к высоким температурам. Кроме того, ПЖ этих двух линий оказалась не только выше других инбредных, но и выше ПЖ контрольной линии. Мы рассматриваем это как аргумент в пользу гипотез о необязательном вредном влиянии повышенной гомозиготности и о возможности избежать инбредную депрессию. Выведенные инбредные линии могут послужить основой для дальнейших исследований причин долгожительства уже на генном уровне.
| Конфликт интересов
Не указан. |
Conflict of Interest
None declared. |
Список литературы / References
- Proshkina E.N. Basic mechanisms of longevity: A case study of Drosophila pro-longevity genes / E.N. Proshkina, M.V. Shaposhnikov, A.F. Sadritdinova et al. // Ageing Res Rev. – 2015. – V. 24. – P. 218-231.
- Nebert D.W. Inbreeding and epigenetics: beneficial as well as deleterious effects / D.W. Nebert, M. Gálvez-Peralta, Z. Shi et al.// Nat Rev Genet. – 2010. – V. 11, №9. – P. 662.
- Булыгин А.А. Получение долгоживущих инбредных линий Drosophila melanogaster и Drosophila funebris / А.А. Булыгин, Е.И. Пастухова // Вестник совета молодых учёных и специалистов Челябинской области. – 2019. – № 2(25). – С. 4-10.
- Shenoi V.N. Evolution of increased adult longevity in Drosophila melanogaster populations selected for adaptation to larval crowding / V.N. Shenoi, S.Z. Ali, N.G. Prasad // J Evol Biol. – 2016. – V. 29, №2. – P. 407-417.
- Vermeulen C.J. Changes in mortality patterns and temperature dependence of lifespan in Drosophila melanogaster caused by inbreeding / C.J. Vermeulen, R. Bijlsma // Heredity. – 2004. – V. 92, №4. – P. 275-281.
- Carazo P. Inbreeding removes sex differences in lifespan in a population of Drosophila melanogaster / P. Carazo, J. Green, I. Sepil et al. // Biol Lett. – 2016. – V. 12, №6.
- Hillesheim Correlated responses in life-history traits to artificial selection for body weight in Drosophila melanogaster / E. Hillesheim, S.C. Stearns // Evolution. – 1992. – V. 46, №3. – P. 745-752.
- Puig O. Understanding Forkhead Box Class O Function: Lessons from Drosophila melanogaster / O. Puig, J. Mattila // Antioxidants & Redox Signaling. – 2011. – V. 14, №4. – P. 635–647.
- Schou M.F. Genome-wide regulatory deterioration impedes adaptive responses to stress in inbred populations of Drosophila melanogaster / M.F. Schou, J. Bechsgaard, J. Muñoz et al. // Evolution. – 2018. Doi: 10.1111/evo.13497
- Rogina B. Indy Mutations and Drosophila Longevity / B. Rogina, S.L. Helfand // Front Genet. – 2013. – V. 4.
- Spencer C.C. Testing an ‘aging gene’ in long-lived drosophila strains: increased longevity depends on sex and genetic background / C.C. Spencer, C.E. Howell, A.R. Wright et al. // Aging Cell. – 2003. – V. 2, №2. – P. 123–130.
- Sujkowski A. Endurance exercise and selective breeding for longevity extend Drosophila healthspan by overlapping mechanisms / A. Sujkowski, B. Bazzell, K. Carpenter et al. // Aging (Albany NY). – 2015. – V. 7, №8. – P. 535-550.
- Archer M.A. Breakdown in correlations during laboratory evolution. II. Selection on stress resistance in Drosophila populations / M.A. Archer, J.P. Phelan, K.A. Beckman et al. // Evolution. – 2003. – V. 57, №3. – P. 536-543.
Список литературы на английском языке / References in English
- Proshkina E.N. Basic mechanisms of longevity: A case study of Drosophila pro-longevity genes / E.N. Proshkina, M.V. Shaposhnikov, A.F. Sadritdinova et al. // Ageing Res Rev. – 2015. – V. 24. – P. 218-231.
- Nebert D.W. Inbreeding and epigenetics: beneficial as well as deleterious effects / D.W. Nebert, M. Gálvez-Peralta, Z. Shi et al.// Nat Rev Genet. – 2010. – V. 11, №9. – P. 662.
- Bulygin A.A. Poluchenie dolgozhivushhih inbrednyh linij Drosophila melanogaster i Drosophila funebris [Achieving longevity by inbreeding in Drosophila melanogaster and Drosophila funebris] / A.A. Bulygin, E.I. Pastukhova // Vestnik soveta molodyh uchjonyh i specialistov Cheljabinskoj oblasti [Bulletin of the council of young scientists and specialists of Chelyabinskaya oblast]. – 2019. – № 2(25). – P. 4-10. [in Russian]
- Shenoi V.N. Evolution of increased adult longevity in Drosophila melanogaster populations selected for adaptation to larval crowding / V.N. Shenoi, S.Z. Ali, N.G. Prasad // J Evol Biol. – 2016. – V. 29, №2. – P. 407-417.
- Vermeulen C.J. Changes in mortality patterns and temperature dependence of lifespan in Drosophila melanogaster caused by inbreeding / C.J. Vermeulen, R. Bijlsma // Heredity. – 2004. – V. 92, №4. – P. 275-281.
- Carazo P. Inbreeding removes sex differences in lifespan in a population of Drosophila melanogaster / P. Carazo, J. Green, I. Sepil et al. // Biol Lett. – 2016. – V. 12, №6.
- Hillesheim E. Correlated responses in life-history traits to artificial selection for body weight in Drosophila melanogaster / E. Hillesheim, S.C. Stearns // Evolution. – 1992. – V. 46, №3. – P. 745-752.
- Puig O. Understanding Forkhead Box Class O Function: Lessons from Drosophila melanogaster / O. Puig, J. Mattila // Antioxidants & Redox Signaling. – 2011. – V. 14, №4. – P. 635–647.
- Schou M.F. Genome-wide regulatory deterioration impedes adaptive responses to stress in inbred populations of Drosophila melanogaster / M.F. Schou, J. Bechsgaard, J. Muñoz et al. // Evolution. – 2018. Doi: 10.1111/evo.13497
- Rogina B. Indy Mutations and Drosophila Longevity / B. Rogina, S.L. Helfand // Front Genet. – 2013. – V. 4.
- Spencer C.C. Testing an ‘aging gene’ in long-lived drosophila strains: increased longevity depends on sex and genetic background / C.C. Spencer, C.E. Howell, A.R. Wright et al. // Aging Cell. – 2003. – V. 2, №2. – P. 123–130.
- Sujkowski A. Endurance exercise and selective breeding for longevity extend Drosophila healthspan by overlapping mechanisms / A. Sujkowski, B. Bazzell, K. Carpenter et al. // Aging (Albany NY). – 2015. – V. 7, №8. – P. 535-550.
- Archer M.A. Breakdown in correlations during laboratory evolution. II. Selection on stress resistance in Drosophila populations / M.A. Archer, J.P. Phelan, K.A. Beckman and others // Evolution. – 2003. – V. 57, №3. – P. 536-543.