СЦЕПЛЕННОЕ С ГЕНАМИ ОКРАСКИ ПЕРИКАРПА РИСА НАСЛЕДОВАНИЕ
КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ
Научная статья
Костылев П.И. 1, *, Балюкова Э.С.2, Серапионов Г.А.3
1 ORCID: 0000-0002-4371-6848;
2 ORCID: 0000-0001-8976-2703;
1, 2 Аграрный научный центр «Донской», Зерноград, Россия;
3 Азово-Черноморский инженерный институт Донской ГАУ, г. Зерноград, Россия
* Корреспондирующий автор (p-kostylev[at]mail.ru)
Аннотация
В статье представлены результаты изучения наследования ряда признаков у гибридов второго поколения от скрещивания белозерного сорта риса Кубояр с чернозерным сортом Гагат. Исследования были проведены в АНЦ «Донской» в 2018-2019 гг. Генетический анализ гибридов F2 показал, что расщепление по окраске перикарпа проходило по тригибридной схеме в соотношении 27 черных: 21 коричневых: 16 белых. Наследование высоты растений у F2 происходило по типу частичного доминирования больших значений признака и аллельных различий трех пар генов. По длине метелки – сверхдоминирование больших значений и положительная трансгрессия. По числу колосков на метелке – сверхдоминирование и взаимодействие трех пар генов. По массе 1000 зерен – отрицательное сверхдоминирование признака и дигенные различия родительских форм. По длине зерновки доминирование отсутствовало, родительские формы различались по двум парам генов. По ширине зерен доминировали меньшие значения признака, расщепление происходило по моногибридной схеме. Установлено сцепление локусов всех изученных признаков с генами окраски перикарпа. Проведен отбор выделившихся образцов с черной окраской перикарпа, сочетающих оптимальную высоту растения, длину метелки, озерненность и массу 1000 зерен для селекционной работы.
Ключевые слова: рис, гибрид, наследование, количественные признаки, высота растений, длина метелки, зерновка, окраска перикарпа.
QUANTITATIVE INHERITANCE LINKED TO THE GENES OF THE PERICARP COLORATION OF RICE
Research article
Kostylev P. I. 1, *, Baliukova E. S.2, Serapionov G. A.3
1 ORCID: 0000-0002-4371-6848;
2 ORCID: 0000-0001-8976-2703;
1, 2 Agricultural research center “Donskoy”, Zernograd, Russia;
3 Azov-Black Sea Engineering Institute – affiliate “Don State Agrarian University”, Zernograd, Russia
* Corresponding author (p-kostylev[at]mail.ru)
Abstract
This article presents the results of the research on the inheritance of a number of attributes of the F2-hybrids of the white-grained Kuboyar rice and the black-grained Gagat rice. The study was conducted at the Agricultural research center “Donskoy” in 2018-2019. The genetic analysis of F2-hybrids revealed that pericarp coloration splitting goes according to the following trihybrid scheme: 27 black to 21 brown to 16 white. Height inheritance of the F2 occurred in the form of incomplete dominance of large character values and allelomorphic differences of three gene pairings. On the panicle length, there was superdominance of significant values and positive transgression. On the number of spikelets on a panicle, there was superdominance and interaction of three gene pairings. On the mass of a thousand grains, there was negative superdominance of the character and the digenic differences of the seed parent. On the caryopsis length, there was no dominance, and the seed parents varied in the two gene pairings. On the grain width, there was the dominance of the lesser character values, and the splitting occurred in the monohybrid form. Loci linkage of all the studied characteristics to the pericarp coloration genes was established. The samples having black pericarp coloration with the optimum plant height, panicle length, grains and the weight of a thousand grains were selected for breeding.
Keywords: rice, hybrid, inheritance, quantitative characters, plant height, panicle length, caryopsis, pericarp coloration.
Введение
Рис – это одна важнейшая пищевая культура большинства народов Земли. Обычно используется белый рис, но бывают и сорта с черной, коричневой или красной окраской перикарпа зерна [1].
Эти природные красители (антоцианы и антоцианидины) обладают сильным антиоксидантным и антимутационным действием и очень полезны для здоровья человека [2]. Поэтому селекционеры стремятся создать сорта риса с окрашенным перикарпом и большой урожайностью. Для успеха этой работы нужно знать особенности наследования признаков окраски и продуктивности зерна.
Японские ученые установили, что доминантные аллели в трех локусах Kala1, Kala3 и Kala4 совместно дают черный цвет перикарпа зерна. Коричневые зерна были у растений с доминантным аллелем Kala4, а белые – у растений, несущих рецессивные аллели Kala4, независимо от других генов [3].
Высота растений риса влияет на урожайность за счет устойчивости к полеганию и индекса урожая. Она контролируется локусами количественных признаков (QTL), которые картированы на всех 12-ти хромосомах [4], [5].
Длина метелки риса влияет на такие компоненты урожайности, как количество зерен в метелке, ее плотность и качество зерна риса. Выявлено более 200 локусов количественных признаков длины метелки (QTL), но только небольшое их количество применяется в селекции риса [6].
Количество зерновок на метелке и их масса – это очень важные признаки, непосредственно определяющие урожайность зерна риса. В настоящее время идентифицировано 58 QTL, распределенных по всем 12 рисовым хромосомам, в том числе 22 – по количеству зерен на метелке, 17 – по количеству колосков на метелке и 19 – по фертильности колосков [7]. Susilowati et al. (2017) сообщили о четырех основных QTL для числа зерен на метелке, которые находились в хромосомах 1, 4, 6 и 7 [8].
Высокая урожайность риса в значительной степени зависит от размеров и массы зерновки. Xu F. et al. (2016) исследовали 416 образцов риса с помощью 143 маркеров и обнаружили 27 QTL, влияющих на размеры зерна. Широко распространенный локус GS3 тесно связан с длиной, шириной зерновки и соотношением l/b [9]. Основной QTL для массы зерновки был идентифицирован возле центромеры хромосомы 3, другие QTL – в хромосомах 2, 5, 8 и 12 [10]. Селекция чернозерного риса ведется во ВНИИ риса (Краснодарский край). Выведение такого риса для Ростовской области также является актуальным, поэтому нужно знать особенности наследования признаков у гибридов от скрещивания белозерных и чернозерных сортов.
Цель работы: изучение наследования качественных и количественных признаков у межподвидового гибрида риса Кубояр × Гагат и их сцепления друг с другом.
Материалы и методы
В качестве материала для исследований использовали гибриды F2 из комбинации Кубояр × Гагат. Сорт Кубояр – селекции АНЦ «Донской» среднеспелый, вегетационный период – 125 дней. Высота растений средняя, 85-94 см. Метелка прямостоячая, длиной 15 см, несет 140-170 колосков. Зерна овальные, средней величины, длиной 8,5 мм, шириной 3,5 мм. Масса 1000 зерен – 28-29 г. Перикарп зерновки – белый.
Сорт Гагат – селекции ВНИИ риса, среднепоздний, вегетационный период – 130 дней. Растения высокорослые, 115-120 см. Метелка поникающая, длиной 17-23 см, несет 120-150 колосков в метелке. Сорт длиннозерный, длина зерновки 10,2 мм, ширина – 2,8. Масса 1000 зерен – 31-32 г. Перикарп зерновки – темно-фиолетовый, почти черный.
Исследования проводили в 2018-2019 гг. на полях ОП «Пролетарское» АНЦ «Донской» Ростовской области. Гибридизацию этих сортов провели в 2017 году. Математическую обработку данных проводили с помощью программ Statistica 8 и Excel. Для генетического анализа использовали программу Полиген А.
Результаты исследований
При скрещивании белозерного сорта Кубояр с чернозерным сортом Гагат в F1 формировалось черное зерно, что свидетельствует о полном доминировании этого признака.
Второе поколение расщепилось на три типа окраски, среди которых появилась коричневая. Расщепление проходило по тригибридной схеме в соотношении 27 черные: 21 коричневые: 16 белые. Отклонения фактических частот от теоретических были незначительными. Различия между родительскими сортами заключались в аллельном состоянии трех генов: Kala 1, Kala 3 и Kala 4. У Кубояра они были рецессивными, генная формула – k1k1 k3k3 k4k4, у Гагата – доминантными K1K1 K3K3 K4K4, которые совместно определяли окраску перикарпа зерна. Черная окраска перикарпа формировалась при наличии трех доминантных генов.
Гибридная популяция второго поколения показала широкий спектр изменчивости по количественным признакам растений. Было проведено сравнение конфигураций кривых распределения частот количественных признаков (далее КРЧ) отдельно в каждой из трех групп окраски зерна: черной (ЧЗ), коричневой (КЗ) и белой (БЗ). При отсутствии сцепления с генами окраски эти кривые должны существенно совпадать друг с другом и наоборот, нахождение QTL в одной хромосоме с генами окраски зерна должно повлиять на характер распределения частот признаков по классам.
По признаку «высота растений» родительские формы значительно различались, в среднем на 24,1 см. У сорта Кубояр она составила 91,5 см, у сорта Гагат – 115,6 см, у гибрида в среднем 105,7 см при частичном доминировании больших значений признака (hp = 0,32).
КРЧ гибрида F2 выходили за пределы изменчивости родительских форм, наблюдалось выщепление трансгрессивных форм (рис. 1).
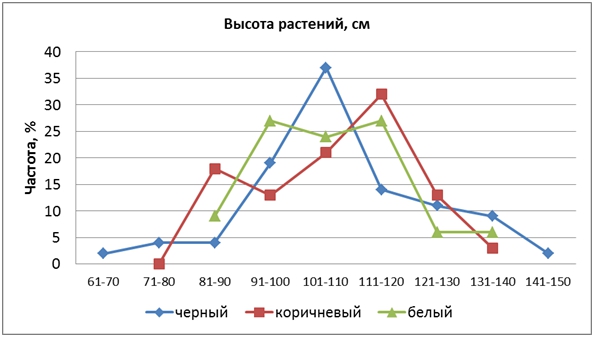
Рис. 1 – Распределение частот признака «высота растений» у гибрида риса
Средние значения высоты в каждой группе окраски различались незначительно, в пределах 105,2-106,8 см. При этом конфигурации их КРЧ были совершенно разными. У форм с коричневыми зернами КРЧ была с двумя вершинами, смещенными влево и вправо от вершины чернозерных форм, при этом правая вершина была больше, чем левая, что свидетельствует о частичном доминировании больших значений признака. КРЧ белозерных форм имела две одинаковые вершины, близко расположенные друг к другу. У чернозерных форм КРЧ была наиболее широкой, охватывая 9 классов, и формировала одну центральную вершину. Большинство растений этой группы имели высоту 101-110 см, однако встречались и карлики (61-70 см) и очень высокорослые (141-150 см). Таким образом, часть генов высоты были расположены в хромосомах 1, 3 и 4, так как показали сцепление с генами Kala.
Длина метелки составила в среднем у Кубояра 14,9 см, Гагата – 19,5 см, гибрида – 20,8 см. Средняя длина метелки составила в группе ЧЗ – 21,0, КЗ – 21,4, БЗ – 19,9 см. Степень доминирования составила hp = 1,36, т.е. наблюдалось сверхдоминирование. КРЧ гибрида имели правостороннюю асимметрию и во всех трех группах окраски выходили вправо за пределы изменчивости большего родительского сорта Гагат, наблюдалась положительная трансгрессия, частота которой составила в среднем 11,7%. При этом КРЧ у форм КЗ была двухвершинной, а у ЧЗ и БЗ – одновершинной. Большая вершина КЗ и вершина БЗ находились в одном классе с величиной признака сорта Гагат (18,1-20 см), а у ЧЗ – смещена вправо (рис. 2).
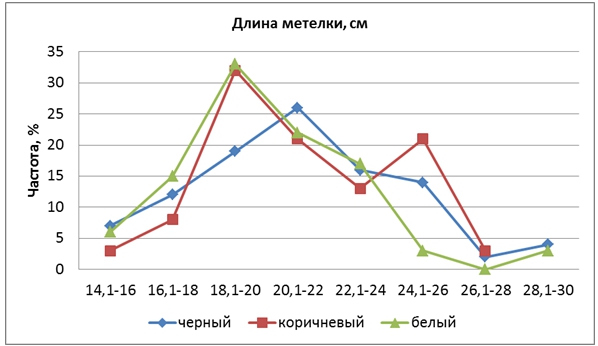
Рис. 2 – Распределение частот признака «длина метелки» у гибрида риса
Вторая вершина форм КЗ находится в классе 24-26 см, и указывает на то, что у форм с коричневым зерном было значительно больше длинных метелок, чем в других группах. Следовательно, гены, контролирующие этот признак, сцеплены с геном Kala 4 в четвертой хромосоме, который обуславливает коричневую окраску перикарпа.
По признаку «число колосков на метелке» исходная родительская форма Гагат (100,2 шт.) уступала сорту Кубояр (141,4 шт.) на 41,2 шт. Среднее количество колосков на метелке у гибрида составило 233,4, варьируя от 44 до 458 штук. Степень доминирования (hp) составила 5,47, что свидетельствует о сверхдоминировании большей величины признака. Установлено взаимодействие трех пар генов, причем у каждой родительской формы были и рецессивные, и доминантные аллели, но в разных локусах. Наблюдалось выщепление множества форм с большим количеством колосков, чем у родительских сортов, что указывает на трансгрессивное расщепление. Среднее число колосков на метелке в группе КЗ (254,3) было значительно больше, чем в ЧЗ (221,0) и БЗ (222,8). Конфигурации КРЧ гибридов в трех группах значительно различались, а их вершины находились в соседних классах (рис. 3).
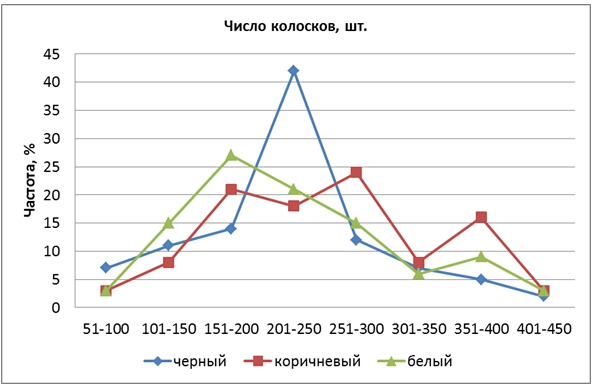
Рис. 3 – Распределение частот признака «число колосков на метелке» у гибрида риса
Левее всех находилась вершина КРЧ гибридных форм БЗ (класс 151-200 шт.), в центре была вершина КРЧ форм ЧЗ (201-250 шт.). У форм КЗ было 3 вершины, две из которых располагались значительно правее остальных. Это указывает на то, что в 4-й хромосоме вместе с геном Kala 4 находятся гены, увеличивающие число колосков на метелке. Для селекции очень значимы положительные трансгрессии, возникающие при рекомбинациях генов.
Общее количество колосков в метелке положительно коррелировало с числом зёрен на ней (r = 0,48±0,08), числом пустых колосков (r = 0,72±0,08) и плотностью метелки (r = 0,92±0,08). Фертильность форм КЗ (48,2%) была значительно выше, чем у ЧЗ (38%) и БЗ (45,4%). Это повлияло и на различия по количеству завязавшихся зерен на метелке: у КЗ – 128,0, у ЧЗ – 83,5, у БЗ – 92,3 штук (в среднем у гибрида 101,3 шт.).
По признаку «масса 1000 зерен» исходные родительские формы различались на 2,8 г. У Кубояра она составляла 28,6 г, у Гагата – 31,4 г. Масса 1000 зерен в F2 варьировала в пределах от 17 до 37 г (в среднем 27,2 г). Расщепление было в соотношении 1:4:6:4:1, что можно объяснить наличием у родительских сортов в двух разных локусах доминантных аллелей с различной силой влияния на величину признака. КРЧ гибридов по группам значительно различались (рис. 4).
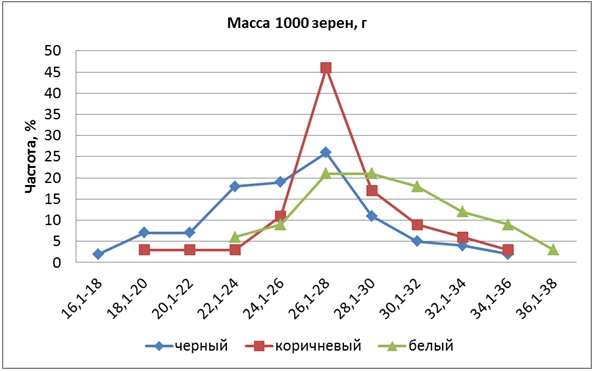
Рис. 4 – Распределение частот признака «масса 1000 зёрен» у гибрида риса
КРЧ форм с БЗ была сдвинута вправо относительно таковой форм КЗ, а у форм ЧЗ, наоборот, влево, что повлияло на среднюю величину этого признака: у форм ЧЗ – 25,8 г, КЗ – 27,8 г, БЗ – 29,2 г. Формы с белым перикарпом имели более крупное зерно, чем с коричневым и особенно с черным. Здесь также проявилось сцепление генов. Выявлено 8 трансгрессивных растений с массой 1000 зерен от 34 до 37 г.
По длине зерновки исходный родительский сорт Гагат (10,5 мм) значительно превосходил сорт Кубояр (8,4 мм) на 2,1 мм. У гибридов этот признак колебался от 7,5 до 12 мм, в среднем – 9,5 мм. Доминирование отсутствовало (hp = 0,05). Расщепление произошло в соотношении 1:4:6:4:1 при взаимодействии двух пар генов.
КРЧ гибрида в трех группах различались по конфигурации и имели по две вершины – большую слева и маленькую справа (рис. 5). Большие вершины в группах КЗ и БЗ находились в одном классе (9,1-9,5 мм), а у форм ЧЗ была смещена вправо и поэтому средняя длина зерновки (9,7 мм) у них была несколько больше, чем у БЗ (9,5 мм) и КЗ (9,4 мм). Это указывает на сцепление локусов длины зерновки с генами окраски перикарпа.
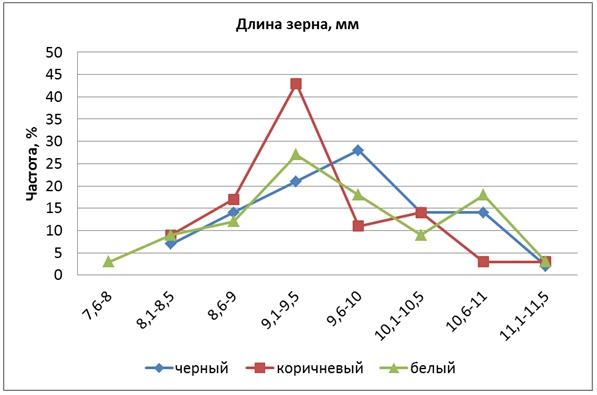
Рис. 5 – Распределение частот признака «длина зерновки» у гибрида риса
По ширине зерен родительские сорта различались на 0,5 мм, у Кубояра она составила 3,4 мм, у Гагата – 2,9 мм. Средняя ширина зерновки у гибридов F2 оказалась 3,07 мм, варьируя в пределах изменчивости родительских сортов от 2,6 до 3,7 мм. Наблюдались правосторонняя асимметрия и доминирование меньших значений признака (hp = -0,32). Расщепление происходило по моногибридной схеме в соотношении 3:1.
КРЧ трех фракций гибрида также различались между собой. Они были одновершинными, причем левая вершина соответствовала формам ЧЗ, правая – БЗ (рис. 6). Посередине между ними располагалась КРЧ форм КЗ.
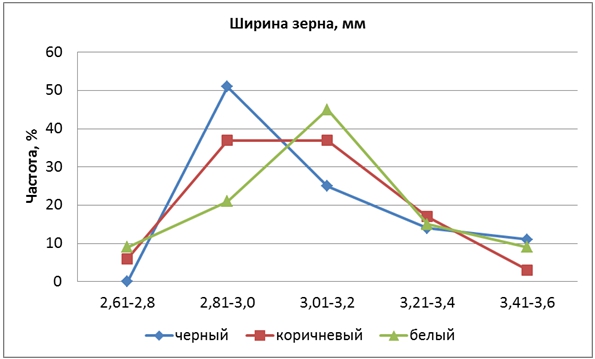
Рис. 6 – Распределение частот признака «ширина зерновки» у гибрида риса
Таким образом, формы ЧЗ имели более длинное, узкое, мелкое зерно по сравнению с другими группами. Для того, чтобы преодолеть это сцепление признаков, нужно искать образцы с новым сочетанием генов, возникающим в результате кроссинговера.
Из популяции F2 были выделены лучшие формы, сочетающие черную окраску перикарпа с оптимальной высотой растений, повышенной озерненностью и массой 1000 зерен. Их высота колебалась в пределах 70-130 см, длина метелки – 14,4-25,0 см, общее число колосков – 98-387 шт., масса 1000 зерен – 20,5-33,3 г, длина зерна – 8,2-10,6 мм, ширина зерна – 2,8-3,5 мм. Эти формы посеяны на третье поколение в гибридном питомнике для последующего создания сортов риса с черным перикарпом зерновки.
Выводы
- Расщепление по окраске перикарпа проходило по тригибридной схеме по типу комплементарности в соотношении 27 черных: 21 коричневых: 16 белых. Черная окраска перикарпа формировалась при наличии в генотипе трех доминантных генов Kala 1, Kala 3 и Kala 4, коричневая – определялась геном Kala 4, белая – остальными комбинациями генов.
- Наследование признака «высота растения» происходило с частичным доминированием больших значений признака (hp = 0,32). Исходные родительские формы различались по аллельному состоянию трех пар генов.
- По длине метелки наблюдали сверхдоминирование больших значений (hp = 1,36) и положительную трансгрессию. Различия между родителями были больше, чем по одной паре генов.
- По числу колосков на метелке установлено сверхдоминирование (hp = 5,47) и взаимодействие трех пар генов.
- По массе 1000 зерен установлены отрицательное сверхдоминирование этого признака (hp = -2,0), расщепление 1:4:6:4:1, т.е. дигенные различия.
- По длине зерновки доминирование отсутствовало (hp = 0,05), расщепление в соотношении 1:4:6:4:1 при взаимодействии двух пар генов.
- По ширине зерен доминировали меньшие значения признака (hp = -0,32). Расщепление было по моногибридной схеме в соотношении 3:1.
- По трем группам форм с черным, коричневым и белым перикарпом конфигурации кривых распределения частот были особенными и не совпадали, что свидетельствует о сцеплении локусов всех изученных признаков с генами окраски перикарпа.
- Для дальнейшей селекционной работы отобраны формы F2 с черной окраской перикарпа, сочетающие оптимальную высоту растения, длину метелки, повышенную озерненность и массу 1000 зерен.
| Конфликт интересов
Не указан. |
Conflict of Interest
None declared. |
Список литературы / References
- Kushwaha U.K.S. Black Rice: Research, History, and Development / U.K.S. Kushwaha // Adv. Plants Agric. Res. 2016. – XX. – 192 p. DOI: 10.15406/apar.2016.05.00165
- Ciulu M. Extraction and Analysis of Phenolic Compounds in Rice: A Review / M. Ciulu, M.L. Cádiz-Gurrea, A. Segura-Carretero // Molecules, 2018. – 23(11). – 2890. https://doi.org/10.3390/molecules23112890
- Maeda H. Genetic dissection of black grain rice by the development of a near isogenic line / H. Maeda, T. Yamaguchi, M. Omoteno, T. Takarada, K. Fujita, K. Murata, Y. Iyama, Y. Kojima, M. Morikawa, H. Ozaki, N. Mukaino, Y. Kidani, T. Ebitani // Breed Sci., 2014. – Vol. 64(2). – P. 134–141. DOI: 10.1270/jsbbs.64.134
- Zhou L. Dissection of genetic architecture of rice plant height and heading date by multiple-strategy-based association studies / L. Zhou, S. Liu, W. Wu, D. Chen, X. Zhan, A. Zhu, Y. Zhang, S. Sheng, L. Cao, X. Lou, H. Xu // Scientific Reports, 2016. – 6. – P. 29718.
- Han Z. QTLs for heading data and plant height under multiple environments in rice / Z. Han, W. Hu, C. Tan, Y. Xing // Genetica, 2017. – 145. – P. 67-77. doi: 10.1007/s10709-016-9946-6.
- Wang X. Identification and application of major quantitative trait loci for panicle length in rice (Oryza sativa) through single‐segment substitution lines / X. Wang, G. Liu, Zh. Wang, S. Chen, Y. Xiao, Ch. Yu // Plant breeding. 2019. – Vol. 138(3). – P. 299-308. https://doi.org/10.1111/pbr.12687
- Niu X. Identification and validation of quantitative trait loci for grain number in rice (Oryza sativa L.) / X. Niu, Y. Zhu, Zh. Sun, S. Yu, J. Zhuang, Y. Fan // Agronomy. 2020. – Vol. 10. – P. 180. doi:10.3390/agronomy10020180
- Susilowati M. Identification of a major quantitative trait locus for grain weight in rice using microsatellite marker / M. Susilowati, H. Aswidinnoor, W. Enggarini, K.R. Trijatmiko // Makara Journal of Science. 2017. – Vol. 21. – No. 4. – P. 155-162. doi: 10.7454/mss.v21i4.6590
- Xu F.F. Association mapping of quantitative trait loci for yield-related agronomic traits in rice (Oryza sativa L.) / F.F. Xu, L. Jin, Y. Huang, Ch. Tong, Y.L. Chen, J.S. Bao // Journal of Integrative Agriculture, 2016. – 15(10). – P. 2192–2202. doi: 10.1016/S2095-3119(15)61244-8
- Yuan H. OsSPL18 controls grain weight and grain number in rice / H. Yuan, P. Qin, L. Hu, S. Zhan, S. Wang, P. Gao, J. Li, M. Jin, Z. Xu, Q. Gao, A. Du, B. Tu, W. Chen, B. Ma, Y. Wang, S. Li // Journal of Genetics and Genomics, 2019. – 46. – P. 41-51. https://doi.org/10.1016/j.jgg.2019.01.003