Для цитирования: Григорян С.С., Исаева Е.И., Бакалов В.В., Осипова Е.А., Бевз А.Ю., Простяков И.В., Надоров С.А. Амиксин – индукция интерферонов альфа, бета, гамма и лямбда в сыворотке крови и легочной ткани // РМЖ. Медицинское обозрение. 2015. №2. С. 93
Введение
Амиксин (тилорон) — низкомолекулярный синтетический индуктор синтеза интерферона (ИФН), относится к классу флуоренонов. Его
Семейство ИФН является ключевым компонентом врожденного и адаптивного иммунитета, обеспечивающего первую линию защиты организма от различных инфекционных агентов [5].
В настоящее время идентифицированы 3 типа ИФН:
1. ИФН
2. ИФН
3. Недавно открытый
Все 3 типа ИФН выполняют роль первой линии защиты организма не только при вирусных, но и бактериальных и паразитарных заболеваниях [15, 16]. Важно отметить, что ИФН продуцируются не только при различной инфекционной патологии, но и под действием ряда высоко- и низкомолекулярных соединений — индукторов ИФН [3, 4].
Известно, что Амиксин стимулирует выработку в организме эндогенных ИФН, которые в ранних исследованиях тестировались биологическим методом — по противовирусному действию — и были отнесены к ИФН -α, -β (
Для оптимизации клинического применения индуктора ИФН Амиксина представлялось целесообразным в рамках доклинического исследования, в строго контролируемых лабораторных условиях, основанных на принципах доказательной медицины, идентифицировать и провести сравнительное изучение индукции препаратом Амиксин ИФН -α (
Цель настоящего исследования — идентифицировать на молекулярном уровне экспериментальные данные по индукции ИФН -α и -β (
Содержание статьи
- 1 Материалы и методы
- 2
Процедура введения препарата
- 2.1 Получение образцов сыворотки и суспензии легочной ткани мышей
- 2.2 Определение ИФН альфа, бета, гамма и лямбда в сыворотке и легочной ткани мышей
- 2.3 Статистическая обработка
- 2.4 Результаты исследований
- 2.5 Индукция интерферона альфа и лямбда в легочной ткани мышей препаратом Амиксин
- 2.6 Сравнительная характеристика динамики продукции интерферонов альфа и лямбда в сыворотке и легочной ткани мышей препаратом Амиксин
- 2.7 Обсуждение результатов исследований
- 2.8 Заключение
Материалы и методы
Дизайн исследований
Исследования проводились в 2 этапа. Первый этап — в Институте органического синтеза Латвии (г. Рига), включал исследование динамики индукции и продукции ИФН -α, -β, -γ и -λ препаратом Амиксин в дозах 40 и 400 мг/кг в сыворотке крови мышей в течение 120 ч.
Второй этап — в лаборатории индукторов интерферона ФГБУ «ФНИЦЭМ им.
Животные
На первом этапе использовались самки 55 мышей линии СВА/СаОlaHsd весом 19–30 г, полученные из вивария Harlan Laboratories, Нидерланды.
На втором этапе — самки 85 мышей линии СВА/CaOlaHsd весом 18–20 г, полученные из питомника «Андреевка» (ФГБУ «НЦБМТ» РАМН).
Содержание животных
Содержание, питание и уход за животными в ходе эксперимента осуществлялись в соответствии с требованиями «Правил проведения работ с использованием экспериментальных животных», Приказ Минздравсоцразвития РФ от 23 августа 2010 г. № 708н. Этические принципы обращения с лабораторными животными соблюдались в соответствии с European Convention for the Protection of Vertebral Animals Used for Experimental and Other Scientific Purposes. CETS No.123. Перед исследованием животные подвергались
Исследуемый препарат
Амиксин® (тилорон) — лекарственная форма: таблетки, покрытые пленочной оболочкой, 125 мг,
Процедура введения препарата
Исследуемый препарат растворяли в стерильной дистиллированной воде в концентрациях, соответствующих вводимым дозам: 20, 40, 400 мг/кг. Экспериментальные животные распределялись на 2 или 3 группы, соответственно этапу исследований и дозам вводимого препарата. Подбор животных в группы осуществлялся методом случайной выборки. Каждая доза препарата Амиксин вводилась мышам однократно перорально (внутрижелудочно) в объеме 50 мкл специальными дозированными зондами размерами, соответствующими физиологической длине пищевода экспериментальных мышей. На каждый срок исследования использовались по 3 мыши.
Получение образцов сыворотки и суспензии легочной ткани мышей
Мышей выдерживали под легким эфирным наркозом до исчезновения ноцицептивных рефлексов. Забор крови проводили методом декапитации в стерильных условиях. Образцы крови, полученные в каждый срок исследования, состояли из пула (смеси) крови от 3 мышей в объеме 3 мл (по 1 мл от каждой особи), из которого после центрифугирования в режиме 5000 об/мин 15 мин отбирали надосадочную сыворотку в объеме 1,0 мл в стерильные пробирки типа Эппендорф. Сыворотки крови мышей, соответствующие каждому сроку забора, разделяли на аликвоты по 0,3 мл в 3 стерильные пробирки Эппендорф, каждую из которых хранили при t -70°C до проведения соответствующего иммуноферментного анализа (ИФА). Для получения суспензии легочной ткани обескровленных мышей фиксировали на специальных парафиновых подложках, в асептических условиях вскрывалась грудная клетка, изолировались и извлекались легкие (правое и левое), которые последовательно трехкратно промывали в буфере Hepes и питательной среде № 199, взвешивали и размельчали сначала в ступке, а затем в специальном гомогенизаторе. Все процедуры проводились в кратчайшие сроки и на холоде. Разрушение клеток ткани легкого достигалось трехкратным замораживанием и оттаиванием. Готовилась 10% суспензия гомогената в питательной среде № 199. Супернатанты суспензии гомогената легочной ткани после низкоскоростного центрифугирования (при 1600 g в течение 30 мин) отбирали и замораживали при -70°С до последующего исследования методом ИФА.
Определение ИФН альфа, бета, гамма и лямбда в сыворотке и легочной ткани мышей
Содержание ИФН -α, -β, -γ и -λ2/3 в пробах сывороток и супернатантов 10% суспензии гомогенатов легочной ткани мышей определялось в 9 временных сроках (0 ч, 4 ч, 8 ч, 12 ч, 24 ч, 48 ч, 72 ч, 96 ч, 120 ч — 3 мыши в каждый срок) после однократного перорального (внутрижелудочного) введения Амиксина в дозах 20, 40, 400 мг/кг.
Количественное содержание ИФН -α, -β, -γ и -λ2/3 (
- Mouse Interferon Alpha ELISA Kit; R & D Systems; США.
- Mouse IFN-β ELISA Kit; PBL Interferon Source; США.
- Mouse IFN-γ ELISA Kit; R&D Systems Europe LTD; Великобритания.
- DuoSet® ELISA; Mouse IL-28A/B (IFN-λ2/3) Kit; R&D Systems; CША.
Расчет оптической плотности исследуемых проб проводили на спектрофотометре — ридере Anthos Labtec c использованием вошера и шейкера (Instruments GmbH, Austria) с последующей обработкой и преобразованием данных оптической плотности в пикограммы в мл и мг (пкг/мл и пкг/ мг) по компьютерной программе Software ADAP. Все исследования проводили в
Статистическая обработка
Результаты подвергались статистической обработке путем расчета среднего арифметического значения (M) и стандартной ошибки к нему (±σ). Оценка статистической значимости различий при межгрупповых сравнениях производилась по двустороннему
Результаты исследований
Индукция интерферонов альфа, бета, гамма и лямбда в сыворотке крови мышей препаратом Амиксин
Однократное пероральное введение Амиксина в дозах 40 и 400 мг/кг вызывало дозозависимую индукцию и продукцию ИФН -α и -λ2/3 в сыворотке крови мышей, максимальное содержание которых через 24 ч после введения препарата в дозе 40 мг/кг составляло 176±1,0 и 398±12,0 пкг/мл, а в дозе 400 мг/кг — 1337±93 и 2231±93 пкг/мл соответственно. Уровень продукции ИФН-λ2/3 в сыворотке крови мышей под действием Амиксина в дозе 40 и 400 мг/кг в этот срок значительно (в 7,5 и 5,6 раза соответственно) превышал таковой ИФН-α (рис. 1).
Сопоставимая динамика определялась в индукции под действием Амиксина и накоплении в крови ИФН -β и -γ, максимальный уровень которых достигался также через 24 ч после однократного введения препарата. Однако количественное содержание ИФН -β и -γ в сыворотке крови экспериментальных животных на пике продукции тестировалось на более низком уровне, чем таковое ИФН-α и -λ2/3. При дозе Амиксина 40 мг/кг максимальный уровень синтеза ИФН-β, как и ИФН-γ, в крови определялся на уровне фоновых терапевтических значений (8,1±4,3 и 4,4±1,2 пкг/мл соответственно). При дозе 400 мг/кг максимальные количества ИФН-β и -γ через 24 ч после введения препарата увеличивались в среднем в 10 раз относительно дозы 40 мг/кг, а относительно контроля — в 110 и 46 раз соответственно (рис. 2).
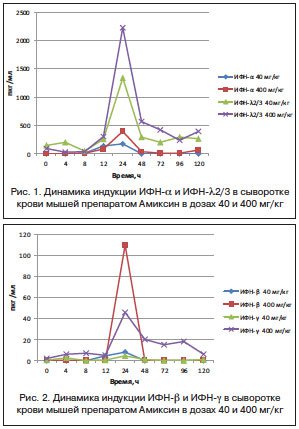
По совокупности результатов, представленных на рисунках 1 и 2, однократное пероральное введение Амиксина в дозах 40 и 400 мг/кг вызывало индукцию в сыворотке крови экспериментальных животных всех типов: -α, -β, -γ, -λ, но количественное содержание ИФН -λ и -α превосходило таковое ИФН-β и -γ в среднем в 10 раз.
Индукция интерферона альфа и лямбда в легочной ткани мышей препаратом Амиксин
По результатам первого этапа исследований были установлены индукция Амиксином всех
Инициация индукции ИФН-α в легочной ткани мышей выражалась в двухкратном повышении через 24 ч после введения Амиксина в дозе 400 мг/кг среднестатистических значений его выработки относительно нулевой точки, а максимальные количества (1125 пкг/мг) синтезировались и аккумулировались только через 48 ч после индукции той же дозой препарата (рис. 3).
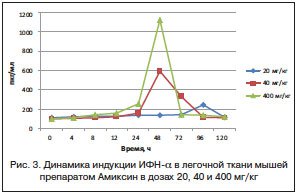
Низкая доза препарата также вызывала двухкратное повышение содержания ИФН-α по сравнению с контролем через 96 ч после его введения. Увеличение дозы вводимого препарата до 40 мг/кг сопровождалось сокращением срока индукции максимальных количеств ИФН-α в легочной ткани мышей к 48 ч (595,0 пкг/мг), причем в течение последующих 24 ч синтез ИФН-α сохранялся на уровне 335,0 пкг/мг.
В целом, под действием различных доз Амиксина в легочной ткани мышей определялась дозозависимая, но более поздняя, относительно таковой в сыворотке крови, продукция ИФН-α (рис. 3).
Амиксин в возрастающих от 20 до 400 мг/кг дозах вызывал также индукцию продукции в легочной ткани мышей ИФН-λ, среднестатистические значения количественного содержания которого на пике продукции составляли от 675,0 до 1547,5 пкг/мг соответственно.
Из данных, представленных на рисунке 4, следует, что через 48 ч после введения Амиксина в низкой дозе — 20 мг/кг — в легочной ткани мышей определялось шестикратное повышение содержания ИФН-λ2/3 относительно контроля в нулевой точке. Увеличение дозы препарата до 40 мг/кг повышало содержание в легочной ткани мышей ИФН-λ2/3 до 1405,0 пкг/мг и сокращало срок индукции его максимальной продукции на 24 ч относительно низкой дозы. Иными словами, двухкратное увеличение дозы однократно вводимого Амиксина приводило к пропорциональному двухкратному повышению количества синтезированного ИФН-λ2/3 и обратно пропорциональному двухкратному сокращению срока достижения пика его продукции. Максимальная доза препарата — 400 мг/кг — смещала пик индукции ИФН-λ2/3 в легочной ткани мышей к 12 ч, а уровень его продукции достигал 1547,5 пкг/мг. При использовании Амиксина в дозах 20 и 40 мг/кг продолжительность синтеза ИФН-λ2/3 относительно нулевой точки и его аккумуляции в легочной ткани экспериментальных животных составляла 96 и 72 ч соответственно. Максимальная доза препарата — 400 мг/мг — вызывала более быстрый (12 ч) и значительный подъем продукции ИФН-λ2/3 (1547,5 пкг/мг), который сохранялся на достаточном уровне (347,5 пкг/мг) относительно контроля до 24 ч включительно.
Таким образом, динамика продукции ИФН-λ2/3 в легочной ткани мышей показала, что увеличение однократной пероральной дозы препарата Амиксин сокращает срок достижения пика продукции ИФН-λ2/3. В дозах 20, 40 и 400 мг/кг максимальный синтез ИФН-λ2/3 в легочной ткани тестируется соответственно через 48, 24 и 12 ч после введения препарата. Причем двухкратное повышение дозы Амиксина с 20 до 40 мг/кг вызывает более чем двухкратное увеличение количества синтезированного в легочной ткани ИФН-λ2/3 на пике его максимальной продукции — с 675,0±5,94 до 1405,0±9,86 пкг/мг соответственно. При последующем
Сравнительная характеристика динамики продукции интерферонов альфа и лямбда в сыворотке и легочной ткани мышей препаратом Амиксин
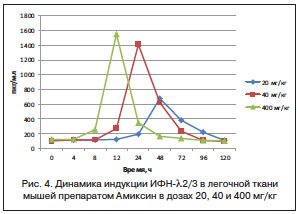
Сравнительный анализ результатов индукции ИФН-α в сыворотке крови и легочной ткани мышей препаратом Амиксин в дозе 400 мг/кг показал значительное количественное преобладание его продукции (в 2,8 раза) в легочной ткани относительно таковой в сыворотке крови экспериментальных животных. Однако пик продукции ИФН-α в сыворотке крови определялся через 24 ч после однократного введения препарата, максимальная продукция ИФН-α в легочной ткани мышей тестировалась только через 48 ч. Иными словами, продукция ИФН-α под действием Амиксина определялась последовательно сначала в сыворотке крови (через 24 ч после введения), а затем в легочной ткани (через 48 ч после введения), но в значительно большем количестве (рис. 5).
Максимальный уровень продукции ИФН-λ2/3 в легочной ткани мышей под действием Амиксина в дозе 400 мг/кг достигался через 12 ч после введения препарата и составлял 1547,5±3,70 пкг/мг, а пик его продукции в сыворотке крови тестировался через 24 ч после введения препарата и составлял 2231±93 пкг/мл, что превышало содержание ИФН-λ2/3 в легочной ткани на пике его продукции в 1,4 раза. Иными словами, продукция ИФН-λ2/3 в легочной ткани опережала по сроку достижения максимального синтеза таковую в сыворотке крови мышей, но определялась на более низком уровне (рис. 6).
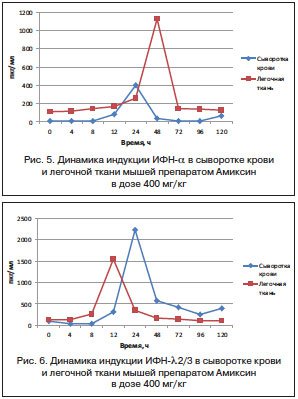
Таким образом, индукция и продукция ИФН-α в сыворотке крови опережает во времени таковую в легочной ткани на 24 ч, но на пике продукции значительно уступает по количественному содержанию (в 2,8 раза). А индукция и продукция ИФН-λ2/3 в легочной ткани опережает по времени таковую в сыворотке крови мышей на 12 ч, но на пике продукции количественно тестируется на более низком уровне, чем в сыворотке крови мышей (в 1,4 раза) (рис. 6).
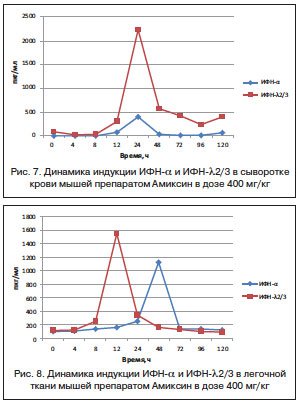
Сравнительный анализ результатов индукции ИФН-α и -λ2/3 в сыворотке крови мышей под действием однократной дозы Амиксина 400 мг/кг показал, что срок достижения их максимальной продукции совпадает и определяется через 24 ч после введения препарата. Однако количественное содержание в крови ИФН-λ2/3 на пике его продукции в 5,6 раза превышает таковое ИФН-α (2231±93 и 398±12 пкг/мл соответственно) (рис. 7).
В легочной ткани мышей максимальный синтез ИФН-λ2/3 под действием Амиксина в той же дозе опережает по времени синтез ИФН-α на 36 ч и превосходит его количественно в 1,4 раза (1547±3,0 и 1125,0±14,43 пкг/мг соответственно) (рис. 8).
Обсуждение результатов исследований
По суммарным результатам проведенных исследований и их сравнительного анализа, препарат Амиксин в дозе 40 мг/кг (эквивалентной суточной дозе для человека) вызывает пролонгированную системную индукцию и продукцию ИФН-α, -β, -γ и -λ в сыворотке крови, максимальный уровень которых определяется уже через 24 ч после его введения, а фоновые значения сохраняются до 120 ч (5 сут) включительно. Одновременно Амиксин в дозе 40 мг/кг индуцирует пролонгированную локальную продукцию ИФН -λ и -α в легочной ткани экспериментальных животных, максимальный уровень которых тестируется соответственно через 24 и 48 ч после введения препарата и также сохраняется в терапевтических значениях до 96 и 120 ч включительно.
Динамика аккумуляции ИФН -α, -β, -γ и -λ в крови при использовании препарата Амиксин в дозе 400 мг/кг (эквивалентной курсовой для людей) повторяет таковую в дозе 40 мг/кг, но количественные показатели ИФН-α и -λ, синтезированных на пике продукции, увеличиваются в среднем в 2 раза, а ИФН-β и -γ — в 10 раз и сохраняются на уровне терапевтических значений в течение 120 ч. Максимальная локальная продукция ИФН-α в легочной ткани мышей под действием Амиксина в дозе 400 мг/кг определяется через 48 ч, а синтез максимальных количеств ИФН-λ2/3 увеличивается и ускоряется на 36 ч, опережая максимальную продукцию ИФН-α в легочной ткани при дозе препарата 400 мг/кг. В последующие сроки исследования — до 120 ч включительно, в легочной ткани экспериментальных животных определяется локальная продукция терапевтических количеств как ИФН-α, так и ИФН-λ.
Таким образом, однократный прием препарата Амиксин в дозе, эквивалентной суточной дозе для человека, в течение первых 24 ч включает синтез ИФН
Многочисленными исследованиями последних лет установлено также, что различные штаммы вируса гриппа, А,
Заключение
В настоящей работе, впервые в строго контролируемых лабораторных условиях, соответствующих принципам доказательной медицины, идентифицированы все известные 3 типа ИФН, которые индуцируются препаратом Амиксин в крови и легочной ткани, подробно изучена динамика, последовательность и профиль их системной и локальной продукции. Согласно результатам проведенных исследований, препарат Амиксин является быстродействующим в первые 24 ч противовирусным средством эндогенной пролонгированной стимуляции выработки ИФН
Таким образом, на сегодняшний день препарат Амиксин является единственным среди зарегистрированных в РФ индукторов ИФН, для которого достоверно доказано действие на выработку всех
Индукция препаратом Амиксин эндогенных ИФН
Литература
- Mayer G.D., Kruger R.F. Tilorone hydrochloride: mode de action // Science. 1970 Vol. 169. P. 1214–1215.
- Чижов Н.П., Смольская Т.Т., Байченко П.И. и др. Клинические исследования переносимости и интерферон-индуцирующей активности амиксина // Вопросы вирусологии. 1990. № 5. С. 411–414.
- Тазулахова Э.Б. Закономерности индукции и продукции α-, β-, γ- интерферона: автореф. дисс. д.б.н. М., 1986.
- Интерферон – 2011: Сборник научных статей. М., 2012.
- Shapira S.D., Hacohen N. Systems biology approaches to dissect mammalian innate immunity // Curr. Opin. Immunol. 2011. Vol. 23. P. 2373–2377.
- Hertzog P.J. Type 1 interferons as primers, activators and inhibitors of innate and adaptive immune responses // Immunol. Cell Biol. 2012. Vol. 90. P. 471–473.
- Hervas-Stubs S., Perez-Garcia J.L. et al. Direct effects of type 1 interferons on cеlls of immune system // Clin. Cancer Res. 2011. Vol. 17. P. 2619–2627.
- Choubey D., Moudgi K.D. Interferons in autoimmune and inflammatory diseases: regulation and roles // J. Interferon &Cytokine Research. 2011. Vol. 12. P. 857–865.
- Hillyer Ph., Mane V.P. et al. Expression profiles of human interferon-alpha and interferon-lambda subtypes are ligand- and cell-dependent // Immunol. Cell Biol. 2012. Vol. 90. P. 774–783.
- Mordstein M., Neugebauer E. et al. Lambda Interferon reders epithelial cells of respiratory and gastrointestinal tracts resistant to viral infections // J. Virol. 2010. Vol. 11. P. 5670–5677.
- Durbin R.K., Kotenko S.V., Durbin J.E. Interferon induction and function at the mucosal surface // Immunol. Rev. 2013. Vol. 255. P. 25–39.
- Jewell N.A., Cline T. et al. Lambda Interferon is the predominant interferon induced by Influenza A virus infection in vivo // J. Virol. 2010. P.11515–11522.
- Donnelly R.P., Kotenko S.V. Interferon-Lambda: A new addition to an old Family // J. Interferon & Cytokine Research. 2010. Vol. 8. P. 555–564.
- Kotenko S.V. IFN –λs // Curr. Opin Immunol. 2011. Vol. 23. P. 1–8.
- Levy D.E., Marie I.J., Induction of type 1 and 3 interferon in response to viral infection // Curr. Opin. Virol. 2011. Vol. 1. P. 476–486.
- Wang B.X., Fish E.N. The yen and yang of viruses and interferons // Trend Immunol. 2012. Vol. 33. P.190–197.
- Григорян С.С., Ершов Ф.И., Поверенный А.М. и др. Особенности продукции интерферона при энтеральном введении индукторов интерферона // Вопросы вирусологии. 1988. №1. C. 67–70.
- Григорян С.С., Иванова А.М., Ершов Ф.И. Противовирусная активность амиксина и его влияние на интерфероновый статус // Вопросы вирусологии. 1990. № 2. C. 138–140.
- Миронов А.Н., Бунатян Н.Д. и др. Руководство по проведению доклинических исследований лекарственных средств. М., 2012.
- Iversen M.B., Paludan S.R. Mechanisms of type 3 interferon expression // J. Interferon & Cytokine Research. 2010. Vol. 8. P. 573–578.
- Onoguchi K., Yoneyama M. et al. Viral infection activate type 1 and 3 interferon gene through a common mechanism // J. Biol.Chem. 2007. Vol. 282. P. 7576–7581.
- Ank N., West H. et al. λ-Interferon ( IFN-λ ), a type 3 IFN, is induced by viruses and IFNs and displays potent antiviral activity against select virus infections in vivo // J. Virol. 2006. Vol. 80. P. 4501–4509.
- Hermant P., Michiels Th. Interferon-λ in the context of viral infections; production, response and therapeutic implication // J. Innate Immune. 2014. Vol. 6. P. 563–574.
- Okabayashi T., Kojima T., Masaki T. et al. Type 3 interferon, not type 1, is predominant interferon induced by respiratory viruses in nasal epithelial cells // Virus Res. 2011. Vol. 1–2. P. 360–366.
- Mordsstein M., Neugebauer E., Ditt V. et al. Lambda interferon renders epithelial cells of respiratory and gastrointestinal tracts resistant to viral infections // J. Virol. 2010. Vol. 84 (11). P. 5670–5677.
- Svetlikova D., Kabat P. et al. Influenza A virus replication is inhibited in IFN-λ2 and IFN-λ3 transfected or stimulated cells // Antiviral. Res. 2010. P. 329–333.
- Khaitov M.R., Laza-Stanca V., Edwards M.R. et al. Respiratory virus induction of alpha-beta- and lambda interferons in bronchial epithelial cells and peripheral blood mononuclear cells // Allergy. 2009. Vol. 63(3). P. 375–386.
- Wang J., Oberley-Deegan R., Wang Sh. et al. Differentiated human alveolar type 2 cells secrete antiviral IL-29 (IFN-λ1) in response to influenza A infection // J. Immunol. 2012. Vol. 182. P. 1296–1304.
- Osterlund P., Pirhonen J., Ikonen N. et al. Pandemic H1N1 Influenza A virus highly sensitive to the antiviral action of interferons // J. Virol. 2010. P. 1414–1422.
- Ank N., Iversen M. et al. An important role for type 3 interferon in TLR-induced antiviral activity // J. Immunol. 2008. Vol. 180. P. 2474–2485.
Поделитесь статьей в социальных сетях
Порекомендуйте статью вашим коллегам
Информация с rmj.ru






